
Если вы не врач, то мы рекомендуем Вам ознакомиться с общей информацией о разрывах ахиллова сухожилия (щелкните мышкой, чтобы перейти к статье для пациентов).
Программу реабилитации после разрывов ахиллова сухожилия вы можете посмотреть здесь.
Узнать о причинах болей в пяточной области и в ахилловом сухожилии при тенопатии ахиллова сухожилия,деформации Хаглунда и при плантарном фасциите («пяточной шпоре») вы можете в отдельных статьях.
Глава 2. АНАТОМИЯ И МЕХАНИЧЕСКИЕ СВОЙСТВА АХИЛЛОВА СУХОЖИЛИЯ
Ахиллово сухожилие (или пяточное сухожилие, tendo calcaneus) – самое крупное и прочное сухожилие в организме человека. Оно образуется в результате слияния сухожилий икроножной (m. gastrocnemius) и камбаловидной (m. soleus) мышц, которые составляют трехглавую мышцу голени (m. triceps surae). Ахиллово сухожилие прикрепляется к пяточному бугру.
Развитое ахиллово сухожилие отсутствует у животных и является отличительной чертой человека. Считается, что оно является одним из главных эволюционных анатомических приобретений, обусловленных прямохождением. Ахиллово сухожилие играло ключевую роль в эволюционном естественном отборе: как у современных человекообразных обезьян, так и у австралопитеков ахиллово сухожилие отсутствует, а появилось оно у представителей рода Homo вероятно более 3 миллионов лет назад [223].
Ахиллово сухожилие выполняет сразу несколько уникальных функций, причем эта многофункциональность подразумевает особую актуальность его повреждений:
1. Прямохождение человека подразумевает положение стопы в анатомической позиции под прямым углом к голени (нейтральное положение). Ахиллово сухожилие тангенциально прикрепляется к самой задней части стопы – к бугру пяточной кости, образуя мощный биомеханический треугольник. Угол между осью большеберцовой кости и линией к пяточной кости у человека один из самых больших среди всех млекопитающих.
2. Мышцы, сухожилия которых образуют ахиллово сухожилие, имеют разные функции и физиологические свойства. M. soleus отвечает за плантарную флексию стопы и в основном содержит медленные мышечные волокна I типа, благодаря которым она функционирует как постуральная мышца, предотвращающая падение человека вперед при стоянии [686]. При этом m. gastrocnemius кроме того отвечает за сгибание конечности в коленном суставе и содержит большее число быстрых мышечных волокон IIB типа, за счет которых возможны энергичные пропульсивные движения, необходимые при быстром беге или прыжках.
3. Так как ахиллово сухожилие прикрепляется к пяточной кости, оно отвечает за движения не только в коленном и голеностопном суставах, но и в подтаранном суставе. Ввиду того, что ось движений в подтаранном суставе обычно проходит выше и медиальнее задне-наружного края пяточной кости [536], ахиллово сухожилие кроме того отвечает и за супинацию стопы [180]. Более того, концентрация сил у медиального и латерального краев энтезиса сухожилия может быть разной.
4. Ротация нижней конечности, происходящая в процессе онтогенеза, приводит к тому, что у взрослого человека ахиллово сухожилие скручено вокруг своей оси: волокна, отходящие от m. gastrocnemius, подходят к латеральной части энтезиса, а волокна, берущие начало от m. soleus, прикрепляются медиальнее [275, 792]. Таким образом, при приложении силы к сухожилию оно способно «раскручиваться», что определяет его амортизационные свойства. Более того, поскольку головки m. gastrocnemius начинаются от надмыщелков бедренной кости, а при сгибании в коленном суставе создается возможность ротационных движений голени, та часть ахиллова сухожилия, которая начинается от m. gastrocnemius может спиралевидно закручиваться вокруг волокон, начинающихся от m. soleus [180]. Этот сложный ротационный механизм дополняется особой формой таранной кости, и вместе эти два биомеханических звена определяют некоторое смещение оси движений в голеностопном суставе по отношению к ахиллову сухожилию во время подошвенного и тыльного сгибания стопы. Смещение оси в свою очередь приводит к небольшой ротации в голеностопном суставе [373].
5. Ахиллово сухожилие обладает широким диапазоном механических свойств. Во время бега ахиллово сухожилие испытывает нагрузки, которые примерно в восемь раз больше, чем вес тела человека, а при стоянии на сухожилие приходится нагрузка вдвое меньше веса тела [430].
Помимо этого, трехглавая мышца голени вместе с ахилловым сухожилием обеспечивает стабильность как в голеностопном, так и в коленном суставах [710].
Макроанатомия
Несмотря на то, что анатомия ахиллова сухожилия широко представлена в работах отечественных и зарубежных анатомов и хирургов, изучавших ультраструктуру и организацию сухожилия [34, 51, 95, 109, 269, 320, 696, 808], структурно-функциональные особенности его скользящего аппарата — паратенона и фасциального футляра [74, 82, 363], интерес морфологов к этому уникальному образованию не ослабевает. Продолжают появляться новые данные о вариантной анатомии мышечно-сухожильного комплекса [210]. Ранее предполагалось, что васкуляризация ахиллова сухожилия изучена полностью [6, 34, 48, 97, 242, 346, 353, 352, 684, 688], однако новые методы регистрации количественных и качественных показателей кровотока также открывают новую страницу в морфологии ахиллова сухожилия [257]. Исследования иннервации и функционального предназначения многочисленных внутрисухожильных рецепторов [28, 74, 131, 134, 696] также не в полной мере позволяют понять принципы проприоцептивных механизмов [520].
Впервые детальное описание образования ахиллова сухожилия за счет слияния сухожилий mm. gastrocnemius было сделано Cummins и соавторами в 1946 году на основе 810 анатомических диссекций [275]. Медиальная (caput mediale) и латеральная (caput laterale) головки икроножной мышцы начинаются от задней поверхности надмыщелков бедренной кости (epicondyli femoris), при этом медиальная головка выражена сильнее [27]. Обе головки соединяются примерно на середине голени и переходят в относительно тонкое сухожилие икроножной мышцы. Так как оно по форме плоское, его еще называют апоневрозом икроножной мышцы. По данным Cummins и соавторов, длина апоневроза варьирует от 12 до 26 см [275]. Волокна икроножной мышцы лежат поверхностнее плоскости апоневроза, который начинается преимущественно от глубоких ее волокон [210]. Несмотря на то, что передняя поверхность икроножного апоневроза и задняя поверхность камбаловидного апоневроза находятся в непосредственной близости, между ними отсутствуют соединительнотканные перемычки. Только в самой нижней части икроножного апоневроза происходит полное соединение (Рисунок 4). Длина икроножного апоневроза вариабельна.
У 2,9%-5,5% людей может присутствовать третья головка икроножной мышцы, в большинстве случаев идущая вместе с медиальной головкой [199]. В некоторых случаях в роли третьей головки икроножной мышцы может выступать непостоянная (отсутствует в 7% случаев по данным 810 анатомических исследований [275]) рудиментарная подошвенная мышца (m. plantaris), брюшко которой может не образовывать собственного сухожилия и конвергировать с медиальной и латеральной головками икроножной мышцы в месте их слияния [199]. Иногда сухожилие подошвенной мышцы вплетается в собственно ахиллово сухожилие [27]. Редко латеральная головка икроножной мышцы может переходить в отдельное сухожилие [199].
Камбаловидная мышца (m. soleus) плоская, почти целиком покрыта икроножной мышцей, лежит глубже нее. Начинается от задней поверхности головки и верхней трети малоберцовой кости и узкой полосой от linea musculi solei, идущей по задней поверхности большеберцовой кости (ниже места прикрепления m. popliteus и выше мест начала m. tibialis posterior и flexor digitorum longus). Вверху с медиальной стороны камбаловидная мышца образует сухожильную дугу (arcus tendineus musculi solei), под которой проходят подколенная вена, артерия и большеберцовый нерв. Ниже мышца также переходит в плоское сухожилие (апоневроз), длина которого варьирует от 3 до 11 см [798]. Далее апоневроз соединяется с апоневрозом икроножной мышцы и образуется ахиллово сухожилие. По сравнению с апоневрозом икроножной мышцы, апоневроз камбаловидной мышцы толще и короче [210, 275]. Волокна камбаловидной мышцы лежат кпереди от плоскости апоневроза, который начинается преимущественно от ее задних волокон (Рисунок 5).
В ряде случаев часть камбаловидной мышцы, начинающаяся от большеберцовой кости (малоберцовая «головка») может отсутствовать, а иногда может иметься дополнительная камбаловидная мышца (m. soleus accessorius), идущая между сухожилием камбаловидной мышцы и длинным сгибателем большого пальца стопы. Добавочная камбаловидная мышца может участвовать в образовании ахиллова сухожилия, а иногда ее сухожилие прикрепляется к пяточной кости независимо, или вплетаться в медиальную коллатеральную связку голеностопного сустава [199]. Обычно по задней поверхности камбаловидная мышца проксимально покрыта икроножной мышцей, а более дистально волокна камбаловидной мышцы выступают с боков от апоневроза икроножной мышцы, что весьма ценно при выполнении селективной электромиографии или биопсии [801].
Обычно соединительнотканные волокна апоневроза камбаловидной мышцы начинаются по задней поверхности мышечных волокон, причем начало апоневроза выше, чем у икроножной мышцы (Рисунок 7H). Таким образом, на уровне соприкосновения мышечных волокон икроножной и камбаловидных мышц последняя разделена сухожильной частью на две мышечных головки (плотная фиброзная ткань по задней поверхности в фронтальной плоскости и тонкая соединительнотканная прослойка в саггитальной плоскости, Рисунок 7H). Аналогичным образом происходит деление m. quadriceps femoris, у которой передняя поверхность m. vastus intermedius представлена апоневрозом, лежащим под m. rectus femoris, но отделена от нее соединительной тканью. Такая организация, вероятно, обеспечивает относительно независимые биомеханические усилия.
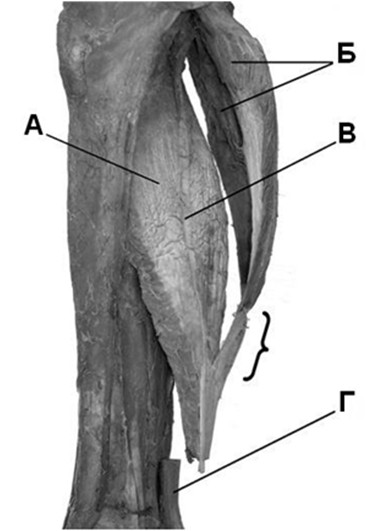
Апоневроз камбаловидной мышцы, идущий по ее задней поверхности, соединяется с апоневрозом икроножной мышцы примерно посередине голени (Рисунок 7). Уровень и геометрия соединения апоневрозов отличаются значительной вариабельностью. Впоследствии общий апоневроз камбаловидной и икроножной мышц идет в дистальном направлении по задней поверхности камбаловидной мышцы, где все еще есть ее мышечные волокна. При этом в общий апоневроз по его ходу продолжают вплетаться дополнительные сухожильные волокна, идущие от камбаловидной мышцы. Кроме того, как мы уже отмечали, в толще камбаловидной мышцы проходит узкое внутримышечное сухожилие, разделяющее ее на две части. Это внутримышечное сухожилие также вплетается в ахиллово сухожилие, но наиболее дистально (Рисунок 7G) [801]. Обычно полная конгломерация апоневрозов происходит в 8-10 см выше пяточной кости.
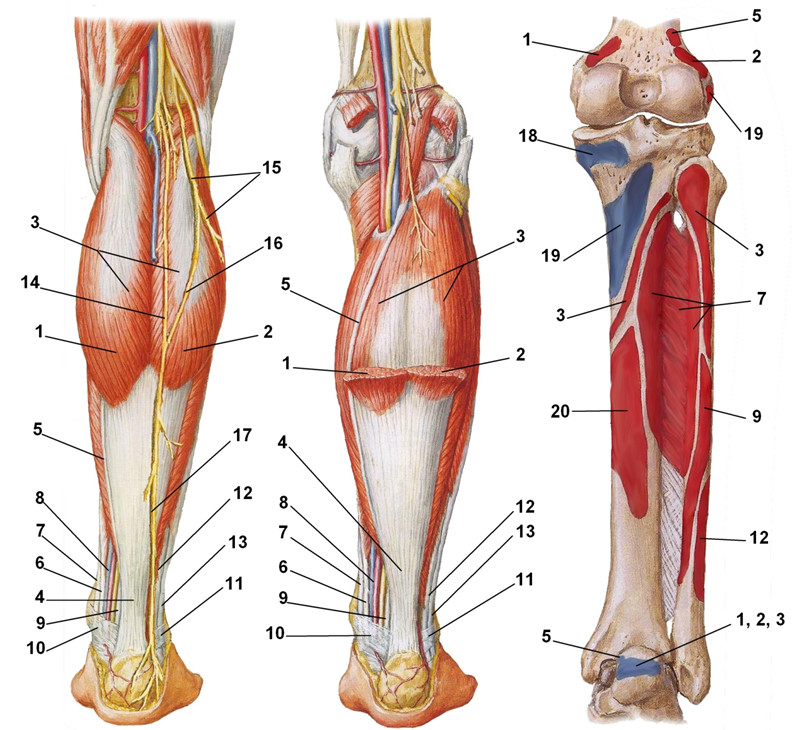
1 – m. gastrocnemius, caput mediale; 2 – m. gastrocnemius, caput laterale; 3 – m. soleus; 4 – tendo calcaneus seu Achilles; 5 – m. plantaris (tendo); 6 – m. flexor digitorum longus (tendo); 7 – m. tibialis posterior (tendo) ; 8 – a., v. tibialis posterior; 9 – m. flexor hallucis longus (tendo); 10 – retinaculum musculorum flexorum; 11 – retinaculum musculorum peroneorum; 12 – m. peroneus brevis (tendo); 13 – m. peroneus longus (tendo); 14 – n. cutaneus surae medialis; 15 – n. cutaneus surae lateralis; 16 – ramus communicans peroneus; 17 – n. suralis; 18 – m. semimembranosus; 19 – m. popliteus; 20 — m. flexor digitorum longus
Принято считать, что ахиллово сухожилие начинается после слияния апоневрозов, однако такое определение не совсем точно отображает вариабельность анатомии. Например, иногда сухожилие камбаловидной мышцы проходит отдельно от сухожилия икроножной мышцы вплоть до бугра пяточной кости, где оно может также и прикрепляться отдельно [559]. Еще одним редким случаем вариантной анатомии является раздельный ход и прикрепление к пяточной кости сухожилий от медиальной и латеральной головок икроножной мышцы (причем эти сухожилия также отделены от сухожилия камбаловидной мышцы) [199]. Такие варианты анатомии при общем осмотре пациента могут привести к ошибочной мысли о патологическом утолщении ахиллова сухожилия на фоне хронического тендинита.
После соединения апоневрозов камбаловидной и икроножной мышц они сужаются и приобретают форму эллипса в поперечном сечении. При этом сухожильные волокна, идущие от икроножной мышцы, закручиваются вокруг волокон от камбаловидной мышцы и в конечном итоге прикрепляются к бугру пяточной кости с латеральной стороны энтезиса. Сухожильные волокна от камбаловидной мышцы идут также не прямолинейно, а косо, и прикрепляются преимущественно с медиальной стороны. Степень ротации сухожильных волокон непостоянна, у некоторых людей волокна от икроножной мышцы закручиваются сильнее и прикрепляются к передней части энтезиса, а у некоторых слабее и прикрепляются к задней части. Впервые вариантность спиралеобразного хода сухожильных волокон отметил еще Cumminis [275], который выделил три типа закручивания волокон (Рисунок 6).
Кроме того, при сильном закручивании волокон проще определить вклад апоневрозов в образование ахиллова сухожилия [180]. Наиболее заметна эта ротация сухожильных волокон в дистальной части сухожилия (в промежутке выше пяточной кости на 5-6 см, Рисунок 7J). Этот спиралеобразный ход сухожильных волокон приводит к тому, что как при расслабленном, так и при напряженном сухожилии его волокна лучше сохраняют пространственную организацию, за счет чего снижается трение между отдельными сухожильными волокнами, и в конечном итоге повышается механическаю прочность сухожилия [138]. Кроме того, это позволяет сухожилию иметь эластичные свойства, удлиняться в соответствующие фазы локомоции. В результате этого процесса внутри сухожилия накапливается энергия, повышающая эффективность икроножно-камбаловидного комплекса. Без этого сокращение только лишь трехглавой мышцы голени имело бы гораздо меньшую биомеханическую эффективность [147].
Вариабельная часть поверхностных волокон ахиллова сухожилия вообще не прикрепляется к пяточной кости. Эти волокна огибают пяточную кость сзади и вплетаются в подошвенный апоневроз, что чаще встречается у молодых [713]. Кроме того, этот факт свидетельствует в пользу теории миофасциальной непрерывности, которая говорит о том, что крайне мало сухожилий в организме изолированно прикрепляются к кости, а большая часть сухожилий различным количеством волокон вплетается в несколько соседних анатомических структур, что приводит к рассеиванию мест концентрации сил [193]. Myers [586] еще больше расширил концепцию миофасциальной непрерывности и даже ввел понятие бесконечной фасциальной «паутины».
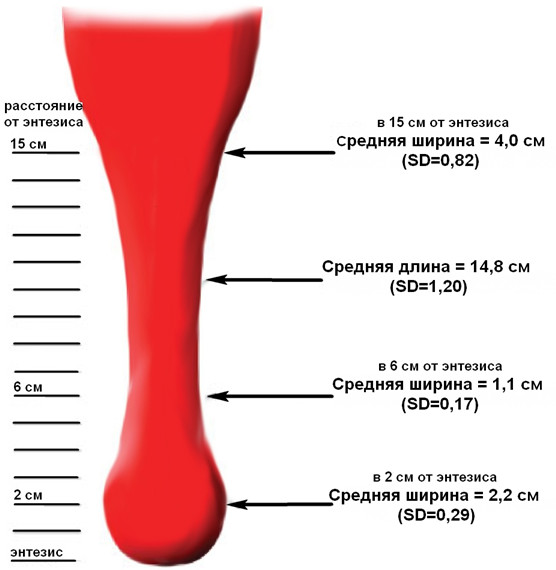
Форма ахиллова сухожилия по его ходу претерпевает значительные изменения (Рисунок 7B–F). Как и большинство других сухожилий в организме, оно распластывается в фронтальной плоскости в области прикрепления к кости. Кроме того, чуть выше места прикрепления оно образует небольшой изгиб кпереди (Рисунок 7F), что видно и при лучевых методах исследования [559]. Обычно в дистальной части толщина сухожилия не превышает 7 мм, а более толстое сухожилие может быть признаком патологического процесса [675]. Собственно в области энтезиса, где сухожилие имеет наиболее плоскую форму в поперечном сечении, его толщина составляет около 3 см, а толщина – 2-3 мм [451]. Площадь поперечного сечения ахиллова сухожилия наименьшая в его средней части, локализованной в 4–7 см проксимальнее места прикрепления к пяточной кости. Среднюю длину ахиллова сухожилия ввиду вариантности анатомии икроножно-камбаловидного апоневроза достаточно трудно определить, но считается, что она составляет 15 см (12-17 см). На поперечных срезах ахиллово сухожилие в средней части овальное и более узкое, чем проксимальная и дистальные части, а проксимальная часть сухожилия на поперечном срезе близка к прямоугольнику (Рисунок 8) [257].
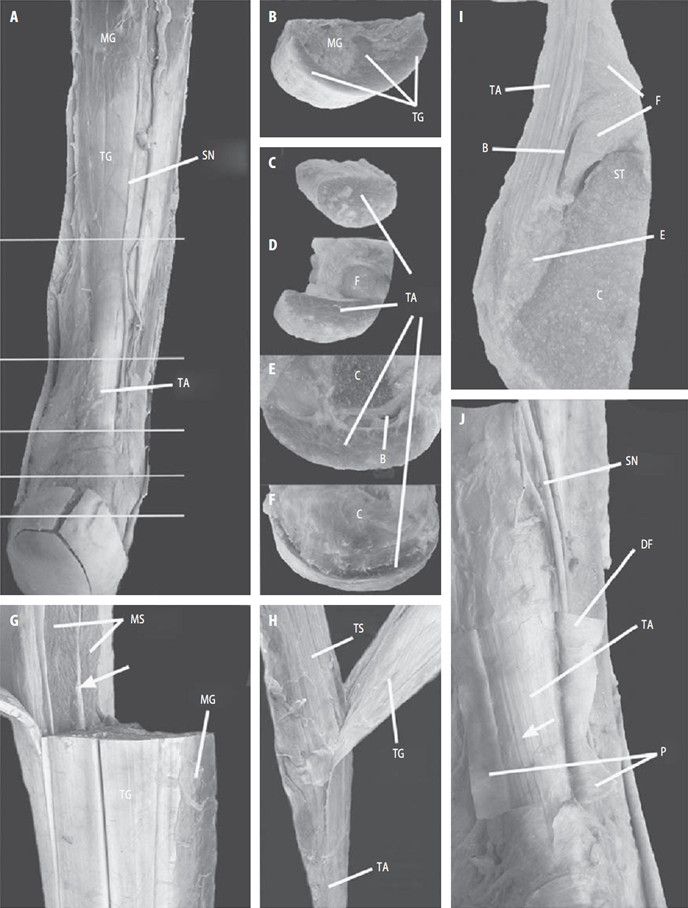
В отличие от других сухожилий, ахиллово сухожилие окружено небольшим количеством истинно синовиального сухожильного влагалища, а преимущественно оно заключено в ложное сухожильное влагалище или паратенон, который образует вокруг сухожилия эластичный рукав, позволяющий ему скользить относительно соседних анатомических структур [403]. Паратенон состоит из нескольких вплотную лежащих друг к другу плотных соединительнотканных мембран, отделяющих сухожилие от глубокой фасции голени. Паратенон имеет густую сеть кровеносных сосудов и хорошо иннервируется. Поверхность самого сухожилия покрыта еще одной мембраной, которую называют эпитеноном. Иногда паратенон и эпитенон называют одним термином – перитенон, который при движениях сухожилия может растягиваться на 2-3 см [588].
Впервые вариантная анатомия икроножного апоневроза была изучена Cummins и соавторами в 1946 году [275]. Наиболее свежие данные принадлежат N. M. Blitz и D. J. Eliot, которые опубликовали результаты своего исследования, основу которого составили 53 анатомических образца, в The Journal of Foot & Ankle Surgery в 2007 году [210].
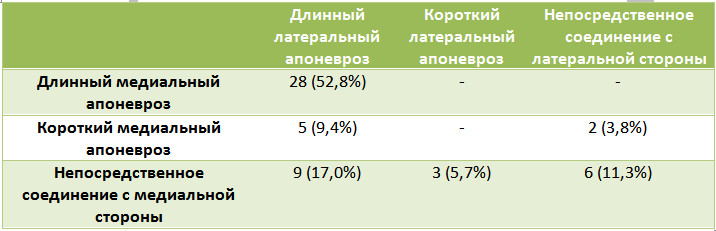
Дистальную границу икроножной мышцы авторы маркировали пятью точками (А, B, С, D, E). Линия, соединяющая эти точки, всегда была похожа на букву «W». Еще одной группой из пяти точек маркировали соединение икроножного и камбаловидного апоневрозов (P, Q, R, S, T). В результате геометрического анализа авторы выделили три основных типа соединения апоневрозов икроножной и камбаловидной мышц: непосредственное соединение, короткий икроножный апоневроз и длинный икроножный апоневроз. Основные варианты анатомии в дальнейшем подразделялись на подтипы в соответствии с длиной медиальной или латеральной стороны икроножного апоневроза (Таблица 1, Рисунок 9).
Таблица 1. Варианты икроножного апоневроза по N. M. Blitz и D. J. Eliot [210].
Вариантная анатомия камбаловидного апоневроза по нашим данным изучалась только W. Pichler и соавторами (2007), которые в результате анализа 80 анатомических образов обнаружили, что в 70% случаев расстояние от наивысшей точки бугра пяточной кости до самых нижних мышечных волокон камбаловидной мышцы находилось в пределах от 2,5 до 7,5 см, в 17,5% — более 7,5 см и в 12,5% случаев – менее 2,5 см [630]. Эти данные звучат несколько неожиданно, поскольку до этого считалось, что ахиллово сухожилие значительно длиннее. Однако наиболее дистальную точку камбаловидного мышечно-сухожильного перехода не стоит считать началом ахиллова сухожилия, поскольку небольшое количество мышечных волокон по передней поверхности апоневроза не нарушают функцию и морфологию истинного сухожилия, а только лишь подчеркивают уникальность ахиллова сухожилия. В любом случае вариантную анатомию как икроножного, так и камбаловидного апоневрозов нужно иметь ввиду, поскольку эти апоневрозы могут использоваться при различных видах сухожильной пластики.
В литературе можно встретить описание и такого редкого варианта анатомии, как непосредственное прикрепление камбаловидной мышцы к пяточной кости кпереди от ахиллова сухожилия, образованного только лишь апоневрозом икроножной мышцы. Впервые такой вариант анатомии был описан J. Bankart и соавторами в 1869 году [178], а в 1999 году он встретился R. Din и G. Matthews [292].
Топографическая анатомия
Непосредственно поверхностнее паратенона лежит глубокая фасция голени (Рисунок 7J), которая сливается с паратеноном рядом с пяточной костью. Функция этой фасции – дополнительная поддержка сухожилия. Кроме того, волокна фасции имеют преимущественно поперечный ход выше пяточной кости, что обуславливает изгиб ахиллова сухожилия кпереди [499, 690]. Благодаря такому ходу волокон фасции и изгибу сухожилия при плантарной флексии сухожилие не натягивается и не выступает подобно тетиве лука. Таким образом, биомеханическую функцию глубокой фасции сложно переоценить, поскольку она делает угол, под которым сухожилие прикрепляется к пяточной кости, относительно постоянным и минимизирует его изменения при движениях стопой.
Икроножный нерв (n. suralis) лежит непосредственно на паратеноне (Рисунок 5A, J) и в большинстве случаев пересекает латеральный край сухожилия в 10-11 см выше энтезиса [262, 788].
Мышечное брюшко подошвенной мышцы (m. plantaris) на дне подколенной ямки проходит вдоль верхнего края латеральной головки икроножной мышцы, а длинное и тонкое сухожилие подошвенной мышцы идет параллельно медиальному краю ахиллова сухожилия. Обычно сухожилие подошвенной мышцы прикрепляется к пяточной кости с медиальной стороны от энтезиса ахиллова сухожилия (в 47% случаев по данным Cummins и соавторов [275]), но в 36,5% из 200 случаев, в которых авторам вообще удавалось идентифицировать сухожилие подошвенной мышцы, оно прикреплялось к пяточной кости слегка кпереди от медиального края энтезиса ахиллова сухожилия [275]. В таких случаях подобный вариант прикрепления подошвенного сухожилия к пяточной кости обеспечивает поддержку передне-внутреннего края bursa retrocalcanea . При третьем варианте, который встречается в 12,5%, сухожилие подошвенной мышцы в дистальной своей части распластывается и вплетается в ахиллово сухожилие по его задней и медиальной поверхности чуть выше энтезиса. Наконец, в 4% случаев сухожилие подошвенной мышцы сливается с ахилловым сухожилием выше места прикрепления последнего к пяточной кости [275].
Возле места прикрепления к пяточной кости ахиллово сухожилие окружено двумя сумками [322]. Одна из них, поверхностная (bursa superficialis tendinis calcanei), лежит между сухожилием и кожей и обеспечивает их взаимное скольжение. Вторая сумка, глубокая (bursa retrocalcanea seu tendini calcanei), располагается между сухожилием, верхним краем бугра пяточной кости и задним краем большеберцовой кости и также способствует свободным движениям сухожилия [798]. В норме пальпация ретрокальканеальной сумки невозможна, при бурсите она пальпируется как мягкоэластичное образование.
В bursa retrocalcanea клинообразно выступает нижний край жировой прослойки Kager, определяемой на рентгенограммах в виде одноименного треугольника. Эта жировая прослойка лежит между m. flexor hallucis longus спереди и ахилловым сухожилием сзади. Относительный размер этой жировой прослойки неодинаков у новорожденных и взрослых [323], однако этот факт не имеет разумного объяснения.
Bursa retrocalcanea имеет сложную форму. Вверху она имеет два язычка, между которыми лежит нижний край жировой прослойки Kager, а в нижней части она полого огибает верхнюю и заднюю поверхности пяточной кости, образуя изгиб кпереди. Контуры здоровой сумки гладкие, а в ее полость может быть введено 1–1.5 мл контрастного вещества. Рентгенологические исследования показали, что она может сообщаться с поверхностной сумкой [323]. На магнитно-резонансных срезах bursa retrocalcanea, в норме содержащая прозрачную вязкую жидкость [237], определяется в виде области сигнала высокой интенсивности [218]. У здоровых людей нижний край жировой прослойки Kager при подошвенном и тыльном сгибании стопы соответственно входит и выходит из bursa retrocalcanea [746]. Возможно, что эта особенность изменяет угол прикрепления сухожилия к кости в зависимости от положения стопы [236]. У симптоматичных пациентов bursa retrocalcanea увеличена, однако, как это ни парадоксально, в нее может быть введено меньшее количество контрастного вещества [322].
Кровоснабжение
Одной из основных патогенетических теорий разрыва ахиллова сухожилия является сосудистая теория. Частота разрыва составляет около от случаев 7 на 100 тысяч населения в год [271, 510], и, как правило, разрыв происходит в 2-7 см выше прикрепления сухожилия к пяточной кости [232, 469, 662]. Ахиллово сухожилие в целом кровоснабжается слабо (в литературе иногда даже можно встретить термин «бледное сухожилие» [257, 623, 746]), а по данным многих авторов, в зоне разрыва васкуляризация сухожилия наименьшая, что и определяет интерес исследователей к кровоснабжению ахиллова сухожилия [138, 242, 468, 688, 723, 820]. Однако в разных исследованиях приводятся неодинаковые данные о локализации разрыва: 2-6 см [469], 4-7 см [662] или 3-5 см [232] выше места прикрепления сухожилия к пяточной кости. Попытки установить корреляцию между кровоснабжением ахиллова сухожилия и локализацией разрыва предпринимались многими авторами [138, 167, 170, 242, 468, 688, 820], но единое мнение на этот счет до сих пор отсутствует [746].
Все сухожилия кровоснабжаются сосудами, отходящими из трех источников: мышечно-сухожильного перехода, окружающей соединительной ткани и области прикрепления сухожилия к кости [549].
Перфузия ахиллова сухожилия осуществляется сосудами паратенона, которые являются ветвями задней большеберцовой артерии (a. tibialis posterior) и малоберцовой артерии (a. peronealis) [138, 242, 418, 688]. Передняя большеберцовая артерия в кровоснабжении ахиллова сухожилия не участвует [138]. Сосуды проникают в толщу сухожилия через структуры, похожие на мезотенон [180]. Сегментарный тип васкуляризации сухожилия предрасполагает к ишемии сегмента сухожилия при хирургическом вмешательстве с выделением сухожилия из окружающих его тканей [121], однако сравнительных исследований на этот счет не проводилось.
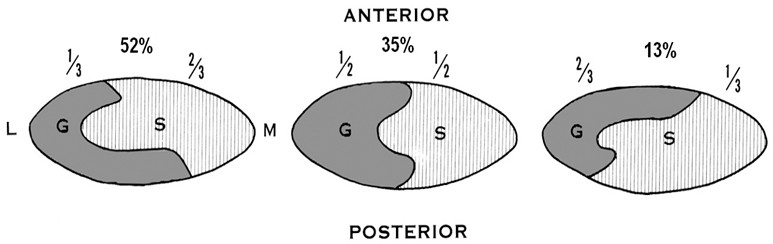
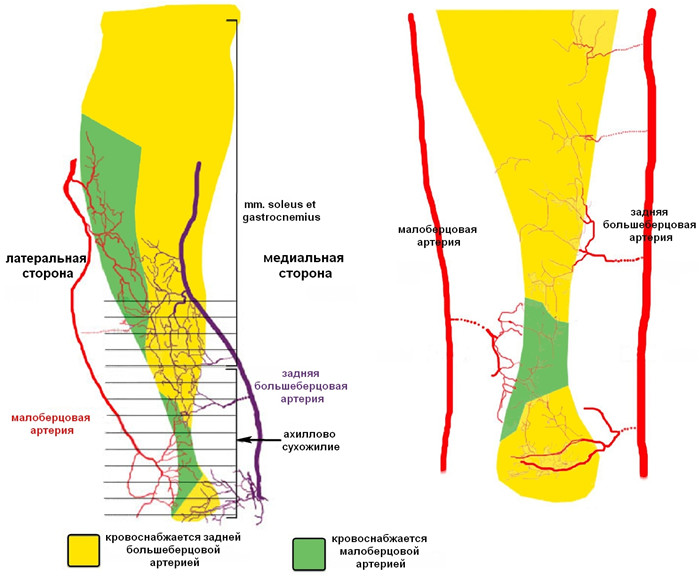
В ранних исследованиях, посвященных изучению васкуляризации ахиллова сухожилия, сообщалось, что оно кровоснабжается равномерно задней большеберцовой артерией и малоберцовой артерией [242, 468, 688, 736, 737]. Позже было установлено, что ахиллово сухожилие разделено на две примерно равные зоны по вертикальной линии: медиальная половина кровоснабжается задней большеберцовой артерией, а латеральная – малоберцовой артерией [173, 738]. В 2009 году T.M. Сhen и соавторы установили, что такое деление не совсем верно, а зоны ответственности артерий за перфузию сухожилия распределяются сложнее (Рисунок 10) [257].
Проксимальная часть сухожилия дополнительно кровоснабжается за счет внутримышечных сосудов, входящих в толщу сухожилия как эндотенон. Однако, большинство авторов считают, что вклад эндотенона в васкуляризацию ахиллова сухожилия невелик [138, 468, 471, 820].
Известно, что паратенон содержит густую сеть кровеносных сосудов [242], однако до сих пор продолжаются дебаты о равномерности их плотности в паратеноне и о более высокой васкуляризации энтезиса [688]. При этом у кроликов паратенон обеспечивает только лишь 35% перфузии пяточного сухожилия, в то время как оставшиеся 65% перфузии обеспечиваются за счет сосудов, проникающих в его толщу из мышц в начале сухожилия и из энтезиса [590]. Отчетливо видно, что вариант кровоснабжения ахиллова сухожилия у человека принципиально иной, с преобладающей ролью паратенона. Вклад сосудов, проникающих из мышцы в проксимальную треть ахиллова сухожилия в составе эндотенона, невелик [138, 468, 471, 488, 684, 820].В дистальной трети ахиллова сухожилия дополнительная перфузия осуществляется мелкими сосудами (диаметр менее 300 µм, [468]) rete arteriosum calcaneare, ветвящимися в надкостнице пяточной кости [301] и являющимися ветвями малоберцовой и задней большеберцовой артерий. Такой вариант дополнительного кровоснабжения сколь-нибудь значительную роль играет только для самых дистальных двух сантиметров ахиллова сухожилия [138, 418, 468, 488, 684, 820]. При этом, сам фибрознохрящевой энтезис в норме не содержит сосудов [170, 192].
С возрастом, в силу ряда причин, абсолютные показатели перфузии ахиллова сухожилия уменьшаются [170, 365]. Помимо этого, с возрастом изменяется и интенсивность перфузии в разных отделах сухожилия. Например, у новорожденных лучше всего из комплекса «трехглавая мышца голени – ахиллово сухожилие – пяточная кость» кровоснабжается энтезис сухожилия, а в возрасте 30 лет наибольшая перфузия сухожилия отмечается в его самой проксимальной части. При этом, вне зависимости от возраста хуже всего кровоснабжается средняя часть ахиллова сухожилия [597].
В литературе можно встретить достаточно разноречивые данные о распределении кровотока в ахилловом сухожилии у взрослых [330]. Ряд авторов считает, что зоной гиповаскулярности является самая дистальная часть сухожилия, непосредственно выше энтезиса [167, 170, 242, 471]. Другие полагают, что хуже всего кровоснабжается средняя часть [428, 468, 723, 820], третьи – дистальная и средняя часть [138, 242], и, наконец, существуют даже исследования, в которых авторы обнаружили локализацию зоны гиповаскулярности в средней и верхней третях сухожилия [688] (Таблица 2).
Таблица 2. Суммарные результаты исследований зон максимального и минимального кровоснабжения ахиллова сухожилия
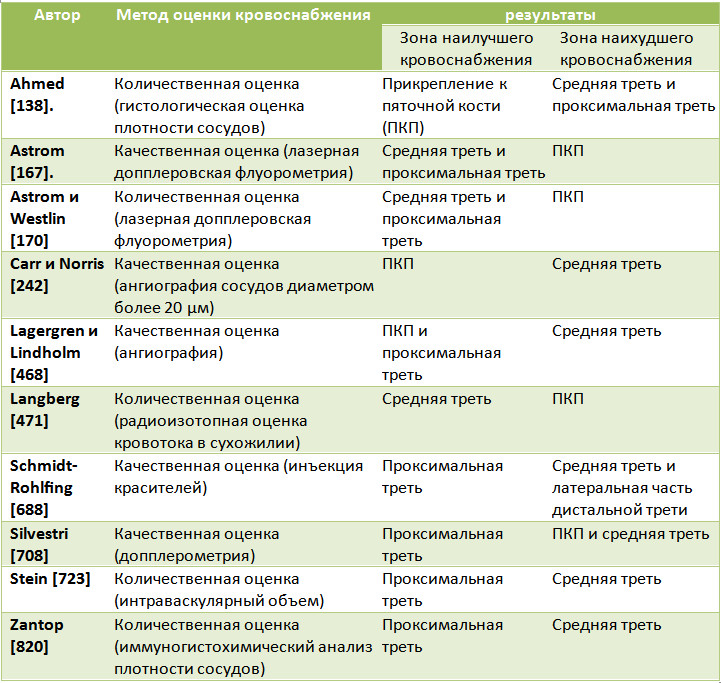
В целом, наиболее распространено мнение о том, что хуже всего ахиллово сухожилие кровоснабжается в его средней части, где перфузия обеспечивается только лишь от сосудов, идущих от паратенона [301, 688]. При этом, распределение кровотока в ахилловом сухожилии может варьировать в зависимости от возраста, пола и физической нагрузки [170].
Иннервация
Сухожилие представляет собой богатейшее рецепторное поле, воспринимающее изменение натяжения мышцы, и является динамическим звеном между костью и мышцей, существующим для эффективного превращения мышечной энергии в движение [121]. Нам не удалось обнаружить полноценных работ, посвященных иннервации ахиллова сухожилия от мышечно-сухожильного перехода до энтезиса. Тем не менее, не стоит отрицать значимость ноцицептивной и проприоцептивной сенсорной иннервации самого сухожилия и его паратенона. Полноценность иннервации ахиллова сухожилия может играть важную роль при его сращении, например периферическая денервация у крыс вдвое снижает способность пяточных сухожилий срастаться после искусственного нарушения их целостности [163].
Сенсорная иннервация ахиллова сухожилия происходит за счет нервов, идущих из мышц (эндотенон) и за счет ветвей соседних кожных нервов, преимущественно икроножного нерва [724]. Паратенон, в отличие от сухожилия, характеризуется более богатой иннервацией и содержит корпускулы Pacinian, которые, предположительно, играют важную роль в механизме проприоцепции [470]. У котов в пяточном сухожилии обнаруживаются комплексы Golgi, локализованные преимущественно в области мышечно-сухожильного перехода и мышечные веретена, располагающиеся более дистально [538].
В пяточном сухожилии крыс существует опиоидная система, которая, вероятно, принимает участие в периферической ингибиции боли [137]. Некоторые сенсорные нервы (волокна типа С) обладают дельта опиоидными рецепторами (ДОР), локализация которых происходит преимущественно в эндотеноне и эпитеноне (где они обычно ассоциированы с кровеносными сосудами) и в паратеноне (меньшая ассоциация с сосудами). Как правило, ДOР связываются с энкефалинами, в результате чего ингибируется ноцицепция и проинфламматорная реакция сенсорных нейропептидов [137]. В норме существует точный баланс между экспрессией опиоидов и сенсорных нейропептидов, который нарушается при патологическом процессе [137]. Существующие знания об иннервации ахиллова сухожилия не позволяют объяснить патогенез боли при тендопатии [309]. Боль в сухожилии может быть обусловлена и изменением сосудистой системы. Наиболее частой гистологической находкой при тендопатии является пролиферация кровеносных сосудов как в толще самого сухожилия, так и в паратеноне [169, 466, 817], а само сухожилие при повреждении может претерпевать ишемические изменения [309].
Структура сухожилия
Биомеханические свойства всех сухожилий обусловлены их молекулярной, клеточной структурой и макроморфологией. Для сухожилий, строение которых соответствует их функции, уместна аксиома всего живого: «Форма определяет функцию, а функция определяет форму». Сухожилия состоят из клеточного и неклеточного компонентов. Клеточный компонент представлен главным образом фибробластами – клетками сложной звездчатой формы, которые синтезируют коллаген и отвечают за реорганизацию экстрацеллюлярного матрикса. Экстрацеллюлярный матрикс состоит из воды (60%–80% общей массы), коллагена (70-86% сухой массы), протеогликанов (1%–5% сухой массы) и эластина (2% сухой массы) [401, 417, 606].
Коллаген, составляющий основу экстрацеллюлярного матрикса, бывает разным. Наиболее распространенный в сухожилиях коллаген I типа образован тремя полипептидными цепями, соединенными ковалентными водородными связями. Молекулы коллагена обладают дополнительными аминокислотными сшивками, которые и обеспечивают механическую прочность. Количество межмолекулярных связей на протяжении сухожилия неодинаково, что определяет разную механическую прочность сухожилия в разных его отделах. Например, наибольшее количество межмолекулярных связей имеется в средней порции сухожилия по сравнению с областями мышечно-сухожильного и сухожильно-костного переходов. При этом средняя часть сухожилия имеет наибольшую механическую прочность, а мышечно-сухожильный переход – наименьшую [608, 757, 807].
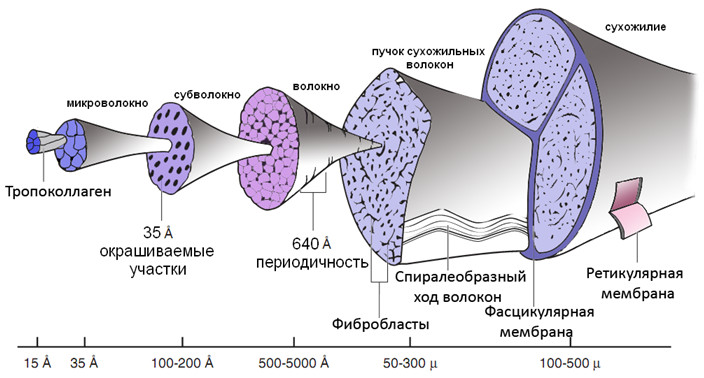
Микроархитектура сухожилия отличается четкой организацией (Рисунок 11). Пять поперечно-связанных молекул коллагена образуют микрофибрилу (микроволокно), которые в свою очередь организуются в субфибрилу (субволокно), а они – в фибриллу (волокно) [421]. Фибриллы, разделяемые протеогликанами, гликозоаминогликанами и водой, располагаются параллельными рядами, и все вместе они образуют экстрацеллюлярный матрикс. Протеогликаны и гликозоаминогликаны, как и коллаген, в высокой степени гидрофильны и состоят из нескольких длинных углеводов, связанных с центральным белком.
Экстрацеллюлярный матрикс вместе с рядами фибробластов образуют собственно сухожильные волокна, которые отделяются друг от друга соединительнотканным эндотеноном, в составе которого проходят кровеносные сосуды и нервы. Эндотенон имеет две мембраны – наружную (ретикулярную) и внутреннюю (фасцикулярную).
В сухожилиях доминирует коллаген I типа (95%), характеризующийся значительной механической прочностью на разрыв [778], которая варьирует от 50 до 100 Н/мм [774, 778]. Однако эта цифра может быть заниженной, так как in vivo коллаген отличается сложной пространственной организацией [429]. Количество эластина в ахилловом сухожилии крайне мало [174, 266]. Эластин может удлиниться в 2 раза перед тем как он разорвется [665]. Если он будет присутствовать в сухожилии в большей пропорции, то трансмиттированная сила к кости будет значительно меньше [657].
Коллаген организуется в гетеротипичные сухожильные волокна, вокруг которых в составе эндотенона располагаются нервы, кровеносные и лимфатические сосуды [667]. В образовании сухожильных волокон также принимают участие коллаген III и V типов [778]. Содержание последних относительно невелико, и они преимущественно регулируют диаметр сухожильного волокна [208]. Исследования показали, что в ахилловом сухожилии пожилых людей преобладают β и γ формы коллагена I типа, что вероятно соответствует процессу увеличения с возрастом связей между волокнами [778].
Хотя нормальное ахиллово сухожилие практически полностью состоит из волокон I типа, при разрывах сухожилия обнаруживается значительное количество коллагена III типа [266]. Культура фибробластов разорванного ахиллова сухожилия синтезирует как коллаген I типа, так и III типа [784]. Коллаген III типа имеет меньшую прочность на разрыв и таким образом его высокое процентное содержание является предрасполагающим фактором спонтанного разрыва.
Коллаген I типа организуется не только в прямые сухожильные волокна, но и в скрученные волокна и в сухожильные пучки, в результате структура сухожилия скорее напоминает многопрядный канат [208]. Каждое отдельное сухожильное волокно не проходит по всей длине сухожилия, так что механическое усилие передается с одного волокна на другое [208]. За эту передачу отвечает аморфный матрикс, в котором располагаются сухожильные волокна. Основу аморфного матрикса составляют коллагеновые волокна VI типа, не организующиеся в волокна, и декорин (богатый лицином протеогликан). Помимо этого, в образовании матрикса ахиллова сухожилия принимают участие фибромодулин, декорин, аггрекан, бигликан, люмикан и версикан [778], которые относительно быстро синтезируются и распадаются [391].
Волокна внутри сухожилия идут волнообразной спиралью, а шаг спирали в среднем равняется 100 μm [208]. Такая геометрия дает волокнам эластичные свойства, при этом существует относительная независимость волокон от экстрацеллюлярного матрикса [208]. Особо важную роль при этом играет эндотенон, который отделяет соседние волокна друг от друга, позволяя им двигаться независимо, и соединяется с эпитеноном на поверхности сухожилия. Эндотенон не содержит соединительной ткани и представлен кровеносными сосудами и нервными волокнами.
Нормальное ахиллово сухожилие, в отличие от разорванного, отличается высокоорганизованным клеточным составом [728]. На поперечном срезе ахиллова сухожилия специализированные фибробласты (теноциты) имеют звездообразную форму, а на продольных срезах они располагаются строгими рядами [667]. Такая строгая клеточная организация является следствием спиралеобразного хода коллагеновых волокон вокруг колонн теноцитов [719] , продуцирующих как волокнистые, так и неволокнистые компоненты экстрацеллюлярного матрикса [637], и возможно определяющих реабсорбцию коллагеновых волокон [207, 688].
У фибробластов есть большое количество плоских отростков, отходящих в стороны от тела клетки к коллагеновым волокнам, благодаря чему коллагеновые волокна закручиваются в завитки. Наиболее длинные и тонкие отростки клеток вытягиваются вдоль ряда фибробластов, при этом отростки соседних фибробластов соприкасаются. Эти межклеточные связи существуют не только в пределах одного продольного ряда фибробластов, но и между соседними рядами. Следовательно, в ахилловом сухожилии имеется трехмерная сеть взаимодействующих отростками клеток, подобная хорошо изученной сети отростков остеоцитов экстрацеллюлярного костного матрикса, которая координирует работу клеток при механической нагрузке. Связи между отростками фибробластов на молекулярном уровне осуществляются за счет коннексина 32 и 43. Коннексин 32 обеспечивает главным образом связи фибробластов внутри одного продольного ряда (т.е. по линии действия основного вектора силы), а коннексин 43 связывает отростки клеток соседних продольных рядов [557]. Waggett и соавторы [777] предположили, что эти различные молекулярные связи играют регулирующую роль в синтезе экстрацеллюлярного матрикса в соответствии с нагрузкой на сухожилие. Оказалось, что коннексин 43 ингибирует синтез коллагена, а коннексин 32 (обеспечивающий связь клеток по вектору силы) наоборот стимулирует.
Энтезис ахиллова сухожилия
Энтезис – место прикрепления любого сухожилия к кости. Он характеризуется особой гистологической структурой, представленной постепенным переходом сухожилия в кость посредством хрящевой зоны (Рисунок 12).
Ахиллово сухожилие прикрепляется к площадке трапециевидной формы по задней поверхности бугра пяточной кости, при этом большая часть области прикрепления находится медиальнее [256]. Средняя высота области прикрепления (расстояние межу верхней и нижней точками энтезиса) 19,8 мм, а средняя ширина – 23,8 мм вверху и 31,2 мм внизу [452]. Таким образом, трапециевидная форма энтезиса обращена вершиной вверх, а сухожилие в дистальном направлении постепенно распластывается по поверхности бугра пяточной кости, что приводит к равномерному распределению механической нагрузки. При этом, несмотря на увеличившуюся площадь сухожилия, не происходит увеличения количества сухожильных волокон. Вокруг сухожильно-костных переходов других сухожилий в состав энтезиса входит жировая муфта, однако энтезис ахиллова сухожилия лишен ее [194].
Как и у других сухожилий, угол, под которым ахиллово сухожилие прикрепляется к кости, относительно постоянен вне зависимости от положения стопы и голени. Когда стопа находится в положении тыльного сгибания, верхняя часть бугра пяточной кости функционирует как направляющий блок для сухожилия, а при плантарной флексии стабилизация положения ахиллова сухожилия обеспечивается за счет глубокой фасции голени (fascia cruris profunda) [180]. При пронации и супинации пяточной кости стабилизирующий механизм менее выражен, хотя и тут глубокая фасция голени, срастающаяся с надкостницей пяточной кости, обеспечивает константность угла прикрепления. Кроме того, угол прикрепления стабилизирует и сам энтезис, который обеспечивает эластическое выравнивание векторов сил [191], сохраняя при этом жесткость соединения, подобно дополнительной оплетке электрического провода у штепселя [689]. Благодаря этому любое физиологическое изменение положения и формы коллагеновых волокон не приводит к концентрации силы в области соединения кости и мягких тканей, а напряжение по экспоненте распределяется на проксимальные отделы сухожилия, что уменьшает риск разрыва в, казалось бы, такой биомеханически уязвимой зоне, как энтезис.
Однако задача распределения нагрузки в области сухожильно-костного перехода ахиллова сухожилия выполняется не только энтезисом. При дорсифлексии стопы передняя поверхность сухожилия плотно соприкасается с верхней частью задней поверхности бугра пяточной кости, что приводит к уменьшению нагрузки на сам энтезис. При оперативном лечении по поводу деформации Haglund резецируют верхний край бугра пяточной кости, что неминуемо изменяет механизм перераспределения нагрузки при соприкосновении передней поверхности сухожилия с костью. Степень увеличения нагрузки на энтезис при этом зависит от объема резецируемой части. Несмотря на то, что точный биомеханический процесс перераспределения нагрузки в зависимости от площади соприкосновения сухожилия и кости не изучен, эту особенность стоит учитывать при планировании объема резекции пяточной кости, особенно у профессиональных спортсменов. При спортивных нагрузках отсутствие соприкосновения передней поверхности сухожилия с верхней частью бугра пяточной кости может привести к катастрофическому повышению нагрузки на энтезис и возможному разрыву в этом редком и весьма неблагоприятном месте.
Перемежающийся контакт сухожилия с верхней поверхностью бугра пяточной кости приводит к локальным специализированным морфологическим изменениям поверхностей этих образований. Верхняя поверхность бугра пяточной кости покрыта толстой фибрознохрящевой надкостницей, а передняя поверхность ахиллова сухожилия в его дистальной части выстлана «сесамовидным фиброхрящем» (Рисунок 12) [671]. Последний термин имеет в своем составе слово «сесамовидный» ввиду того что фибрознохрящевая прослойка располагается не на поверхности сухожилия, а слегка внутри него, подобно сесамовидной кости. Свободным движениям этих двух поверхностей относительно друг друга способствует bursa retrocalcanea , в которую при плантарной флексии вдается языкообразный выступ нижней части жировой подушки Kager. Энтезис, периостальный и сесамовидный фиброзные хрящи, сумка и жировая подушка коллективно функционируют как единый орган, который иногда называют «энтезис-орган» [192, 193]. Эти разнородные ткани объединены одной целью – перераспределить концентрацию сил и снизить риск разрыва в области сухожильно-костного перехода [671, 672].
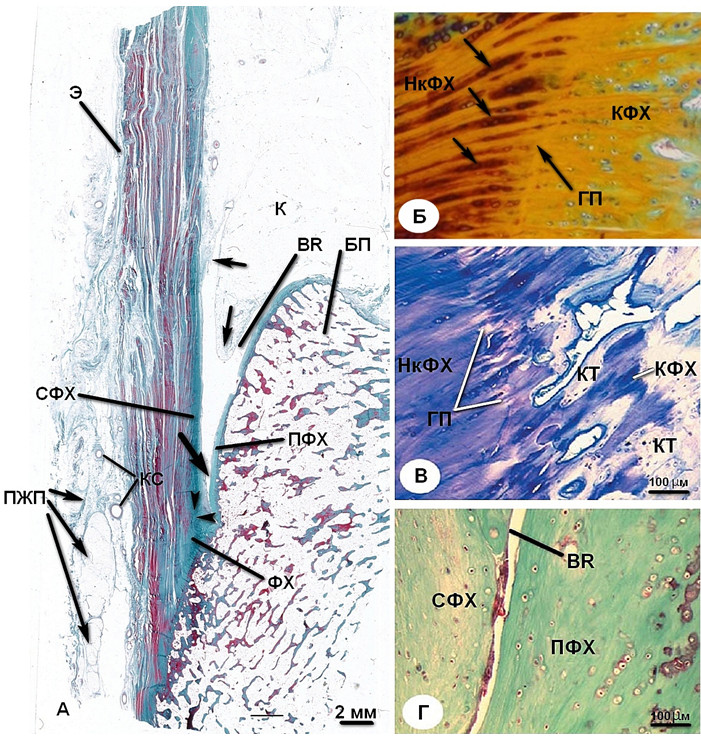
A — Сагиттальный срез, общий вид. Энтезис состоит из выступающего фиброзного хряща (ФХ), который наиболее выражен в верхней части сухожильно-костного перехода (два черных треугольных маркера). Непосредственно проксимальнее сухожильно-костного перехода передняя поверхность сухожилия контактирует (отмечено жирной черной стрелкой) с верхней частью бугра пяточной кости (БП) посредством bursa retrocalcanea (BR). В сумку выступает языкообразный отросток нижней части жировой подушки Kager (K). Проксимальная часть сумки и выступ жировой подушки выстланы синовиальной мембраной (черные стрелки). Дистальная часть сумки с поверхностной стороны образована сесамовидным фиброзным хрящом (СФХ), а с дистальной – периостальным фиброзным хрящом (ПФХ). По задней поверхности ахиллова сухожилия виден эпитенон (Э), содержащий кровеносные сосуды (КС). В самой нижней части ахиллово сухожилие покрыто сзади пяточной жировой подушкой (ПЖП) (трихроматичная окраска по Masson).
Б – сагиттальный срез «границы прилива» (ГП). Клетки фиброзного хряща организованы в продольные ряды (черные стрелки) в зоне некальцифицированного фиброзного хряща (НкФХ). Непосредственно глубже НкФХ располагается кальцифицированный фиброзный хрящ (КФХ) (трихроматичная окраска по Masson).
В – сагиттальный срез глубже «границы прилива». Кальцифицированный фиброзный хрящ постепенно замещается костными трабекулами (КТ) (Окраска толуидином голубым).
Г – сагиттальный срез дистальной части bursa retrocalcanea. По задней поверхности сухожилия располагается сесамовидный фиброзный хрящ (СФХ), а противоположная поверхность кости выстлана периостальным фиброзным хрящом (ПФХ). Синовиальная оболочка на сочленяющихся поверхностях отсутствует (трихроматичная окраска по Masson)
ижениях. Гистологическая граница наоборот не должна быть прямой, чтобы обеспечить прочное якорное соединение сухожилия и кости. Механический парадокс в энтезисе взрослого человека разрешается присутствием на поверхности кости тонкой прослойки кальцифицированного хряща, который можно сравнить с цементом на кирпиче. Присутствие этого слоя обеспечивает гладкий гистологический переход сухожилия (волокна Sharpey) в истинную кость [190].
Волокна Sharpey не являются уникальной особенностью энтезиса ахиллова сухожилия: они обнаруживаются и в других фибрознохрящевых энтезисах, что соответствует как процессу формирования энтезиса, так и малому количеству компактного костного вещества в субхондральной пластинке. У крыс энтезис пяточного сухожилия развивается путем метаплазии фибробластов в плотной фиброзной соединительной ткани сухожилия рядом с поверхностью кости. При этом клетки фиброзного хряща, так же как и их предшественники, фибробласты, организованы в продольные ряды. Вероятно, развитие фиброзного хряща индуцируется механическими стимулами сразу после рождения. При этом ткань функционирует как «мини зона роста» кости. C одной стороны энтезиса фибробласты сухожилия превращаются в клетки фиброзного хряща (граница между зонами плотной фиброзной соединительной ткани и некальцифицированного фиброзного хряща), а с другой стороны кость замещает фиброзный хрящ аналогично эндохондральной оссификации зоны роста длинных трубчатых костей [331].
Фиброзный хрящ энтезиса распространен по плоскости сухожильно-костного перехода неравномерно. Более он выражен в верхней части (т.е. под глубокими слоями ахиллова сухожилия), чем в нижней, где энтезис отличается большим количеством плотной фиброзной соединительной ткани. Примечательно, что именно в задне-нижней части бугра пяточной кости образуются костные шпоры при энтезопатии. Трапециевидная форма фиброзного хряща энтезиса ахиллова сухожилия вероятно способствует функционированию этого образования в качестве мягкотканого направляющего блока благодаря вязкоупругим свойствам [567]. В свою очередь эта функция дополняет более явный направляющий блок, сформированный верхней поверхностью бугра пяточной кости. Однако мягкотканый блок может лишь несильно компенсировать векторы сил и только при коротком плече силы ахиллова сухожилия, например, при дорсифлексии стопы. Quigley и Chaffin [651] вычислили, что расстояние от ахиллова сухожилия до оси движений в голеностопном суставе (плечо силы) уменьшается на 40% при 35 градусах дорсифлексии. Это означает, что наибольшие энергозатраты необходимы для приподнимания веса тела на носках стоп, и именно при дорсифлексии нагрузка на мышечно-сухожильный переход и на само сухожилие максимальна.
В литературе уделяется незаслуженно мало внимания и той части пяточной кости, которая располагается непосредственно под энтезисом. Ее отличительной особенностью является то, что она лишена кортикального слоя. Кость в этом месте имеет хорошо организованные трабекулы, которые преимущественно ориентированы вдоль волокон ахиллова сухожилия, связывая энтезисы сухожилия и подошвенного апоневроза [567]. Архитектоника трабекул предполагает распределение сил по линиям основной направленности внутри кости, соединяя при этом две мягкотканые структуры (ахиллово сухожилие и подошвенный апоневроз). У молодых людей трансмиссия силы происходит не только по трабекулам кости, но и по мягкотканым структурам. Как было отмечено выше, часть волокон ахиллова сухожилия вплетается в подошвенный апоневроз [713]. Аналогичный биомеханический механизм можно наблюдать и в надколеннике – где передача силы частично происходит по костным трабекулам, ориентированным параллельно сухожилию, а частично – по самим сухожильным волокнам [763]. В обоих случаях этот механизм иллюстрирует принцип миофасциальной непрерывности одноименной теории [586].
Биомеханика сухожилия
Экспериментальные исследования прочности мышечно-сухожильно-костного комплекса на конечностях трупов показали, что слабейшим местом в этой системе обычно была мышца [556, 796]. При изолированном исследовании биомеханических характеристик ахиллова сухожилия in vitro (без мышечно-сухожильного перехода) Wren и соавторы обнаружили, что средняя прочность на разрыв превышает 5000 Н, эластичность в области сухожильно-костного перехода составляет 12,5%, а эластичность самого сухожилия – 7,5% [810].
Однако следует подчеркнуть, что измерение истинной прочности мышцы может быть получено только на такой экспериментальной модели, при которой сохранялись бы нервно-рефлекторные связи мышечно-сухожильного комплекса с организмом. Так, при мышечной стимуляции у живых крыс удавалось вызвать разрыв самого сухожилия [180]. В теноцитах присутствуют актин и миозин [390], в результате чего само по себе сухожилие обладает активным контракционно-релаксационным механизмом, регулирующим трансмиссию силы от мышцы к кости [326]. Большинство проводившихся исследований абсолютных цифр прочности на разрыв не учитывали эти особенности [97].
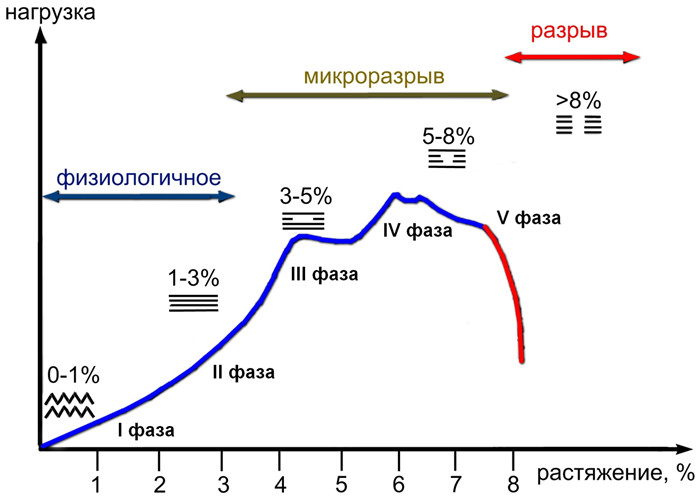
Под действием внешних сил всякое твердое тело деформируется. В теории упругости под деформацией понимают увеличение длины, выраженное в процентах от первоначальной длины образца. Отношение нагрузки к площади поперечного сечения образца называют напряжением. Характер деформации плотной волокнистой соединительной ткани (сухожилие, связка) отражает определенной формы кривая «нагрузка-растяжение» или «напряжение-деформация» (Рисунок 13).
Кривая «нагрузка-растяжение» для ахиллова сухожилия имеет нелинейную форму, что обусловлено спиралеобразным ходом сухожильных волокон и изгибом самого сухожилия [606]. Многие исследователи выделяют до 5 фаз этой кривой [180, 269, 320, 415, 440, 796]:
I фаза. Первоначальное удлинение на 1-2% происходит уже при небольшом напряжении за счет выпрямления извитых коллагеновых волокон.
II фаза. Исчерпаны эластические свойства спиралеобразной деформации сухожильных волокон, которые теперь ориентированы по вектору приложенной силы. Происходит быстрый рост жесткости ткани. Появляющиеся микроразрывы небольшого количества межволоконных связей все еще позволяют восстановить сухожилию первоначальную длину при прекращении силы (эластическое удлинение). Зависимость растяжения от нагрузки линейная.
III фаза. При растяжении сухожилия до 7-8% (точка текучести) происходит прогрессирующее повреждение волокон, но макроскопически ткань еще имеет нормальный вид. Зависимость становится нелинейной, т.е. растяжение возрастает без увеличения деформирующей силы.
IV фаза. Достигается максимальная нагрузка, которую сухожилие может выдержать — предел прочности на разрыв.
V фаза. Полное разрушение пучков сухожилия. Ткань уже не способна поддерживать напряжение.
I фаза кривой «нагрузка-растяжение», при которой коллагеновые волокна начинают выстраиваться линейно, может распространяться до 4%, что соответствует диапазону большинства физиологических нагрузок [606].
Начиная с III фазы, происходит пластическое удлинение сухожилия, достигающее 7-8% (или даже до 16%) от первоначальной длины [97, 180, 320, 602]. При этом прекращение нагрузки не приводит уже к полному восстановлению длины и структуры сухожилия. Эластичность сухожилия, вероятно, обусловлена спиралевидной организацией коллагеновых волокон. Первый нелинейный участок кривой, где наблюдается большое растяжение при малом увеличении напряжения, соответствует выпрямлению извитых волокон, что играет значительную роль в амортизации потенциально опасных ударных нагрузок между мышцей и костью [818]. Если же сухожилие предварительно натянуто, то оно ведет себя как жесткий материал (II фаза кривой). При этом малое смещение сухожилия мышцей передает большое напряжение кости, вызывая немедленный эффект движения. Функция сухожилия определяется особенностями формы кривой «нагрузка-растяжение» лишь частично [247, 368].
Fukashiro и соавторы зарегистрировали максимальную силу, которая трансмиттировалась ахилловым сухожилием при прыжках (in vivo): она составила 2233Н [325, 326]. Komi и соавторы [454], используя специальные измерительные устройства стременного типа, выяснили, что при ходьбе сила формируется и регистрируется непосредственно перед касанием пятки пола, затем в течении 10-20 мсек сила снижается (фаза раннего толчка, удара). Затем сила относительно быстро возрастает и достигает пика в конце фазы отталкивания. Аналогичная закономерность прослеживается при беге. Позже Arndt и соавторы [156] обнаружили, что ахиллово сухожилие подвержено неодинаковым стрессовым нагрузкам в зависимости от модификаций мышечного вклада. Следовательно, разрыв ахиллова сухожилия может произойти вследствие асинхронного сокращения различных компонентов трехглавой мышцы голени или некоординированного сокращения мышц агонистов и антагонистов при нарушенной проприоцепции [513].
С возрастом эластичность и прочность коллагеновых тканей постепенно начинает уменьшаться [602]. Например, если в 25 лет разрывная нагрузка ахиллова сухожилия человека составляет 530 кг, то в 61 год-268, т.е. в 2 раза меньше [97]. Важным фактором, влияющим на прочность сухожилия, является уровень двигательной активности. Иммобилизация и отсутствие нагрузки сухожилия неизбежно влечет за собой значительное уменьшение жесткости и прочности к механической нагрузке. Обездвиживание конечности уже в течение 4-8 недель вызывало заметные адаптивные изменения микроструктуры сухожильной ткани, уменьшение диаметра и площади поверхности коллагеновых волокон [591], снижение активности ферментов биосинтеза коллагена, дегидрогеназ и повышение активности лизосомальных ферментов, а также уменьшение объема кровеносных капилляров в сухожилии [206, 329, 681].
Некоторые аспекты биомеханики и физиологии трехглавой мышцы голени
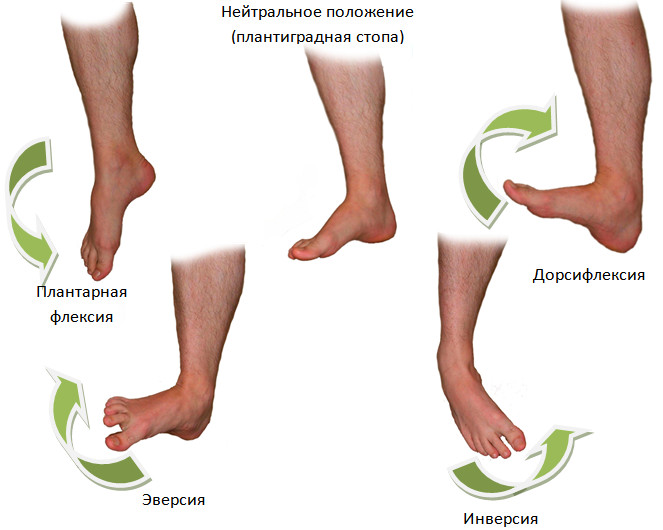
Основная функция трехглавой мышцы голени — подошвенное сгибание стопы. Так как ахиллово сухожилие прикрепляется к пяточной кости, трехглавая мышца голени осуществляет движения не только в коленном и голеностопном суставах, но и в подтаранном суставе. Ввиду того, что ось движений в подтаранном суставе обычно проходит выше и медиальнее задне-наружного края пяточной кости [536], ахиллово сухожилие кроме того отвечает и за супинацию стопы в сочетании с аддукцией (инверсия стопы, Рисунок 14) [180].
Икроножная мышца, являясь двусуставной, одновременно сгибает и голень в коленном суставе. Важной особенностью подобных мышц является способность их отдельных (проксимальных или дистальных) участков при сокращении и растяжении функционировать раздельно. Причиной этого феномена считают независимую иннервацию этих сегментов, а также наличие дополнительной опоры на фасцию и подлежащие мышцы [23, 107].
Камбаловидная мышца оказывает действие только на голеностопный и подтаранный суставы. При обычном для человека типе стояния линия центра тяжести смещена кпереди от оси голеностопного сустава. Сокращенная камбаловидная мышца препятствует тыльному разгибанию стопы под действием веса тела, одновременно формируя свод, который служит опорой для икроножной мышцы, производящей сгибание в коленном суставе. Таким образом, содружественная деятельность этих мышц обеспечивает неотъемлемую человеческую особенность – прямохождение [91]. Сила трехглавой мышцы варьирует от 200 до 500 Кгс [180].
Установлено, что при ритмических сокращениях и растяжении скелетные мышцы функционируют как присасывающе-нагнетательные микронасосы. В основе этой деятельности лежит вибрационный механизм, заключающийся в том, что асинхронно сокращающиеся мышечные волокна, воздействуя на внутримышечные сосуды, способствуют продвижению крови. В условиях гипокинезии микронасосы могут угнетаться, что приводит к ухудшению снабжения мышцы питательными веществами и кислородом [8]. Длительная перегрузка способствует задержке внеклеточной жидкости и повышению напряжения в фасциальном футляре, что может вызывать ишемические боли в мышце и сухожилии [438].
Функциональной единицей мышцы является двигательная единица [ДЕ], состоящая из одного мотонейрона, его аксона и иннервируемых им мышечных волокон. Двигательные единицы мышц разделяют на 2 основных типа. Медленные ДЕ (1-й тип) включают медленный мотонейрон, который иннервирует медленные мышечные волокна, быстрые ДЕ (2-й тип) — соответственно быстрые мотонейроны и быстрые мышечные волокна. Разграничение мышечных волокон прежде всего зависит от активности миозин АТФ-азы. Быстрые мышечные волокна с высокой активностью фермента отличаются более высокой скоростью сокращения. Медленные волокна, в свою очередь, отличаются значительно большей выносливостью. Мышечные волокна подразделяются на 2 подтипа. Подтип 2А занимает как бы промежуточное место между медленными волокнами 1-го типа и быстрыми подтипа 2Б: с одной стороны, они приспособлены к аэробному окислительному способу энергопродукции и поэтому обладают повышенной выносливостью, с другой стороны, располагают мощной лактацидной анаэробной системой энергообеспечения, в связи с чем имеют высокую скорость сокращения. В зависимости от концентрации миоглобина, разные типы волокон отличаются по окраске: медленные — красные, а быстрые — белые [10, 33, 80].
Любая скелетная мышца содержит и быстрые, и медленные волокна, но их соотношение варьирует от функционального предназначения данной мышцы, возраста, пола, индивидуальных особенностей двигательной активности человека. Быстрые волокна преобладают в мышцах, которым требуется точность движений, большая скорость сокращения и максимальная сила, развиваемая в короткий промежуток времени (мышцы кисти и глаз). Медленные же волокна превалируют в мышцах, предназначенных к выполнению длительной работы со стабильным, но невысоким усилием. К таковым относятся мышцы, обеспечивающие поддержание определенного положения тела и противодействие гравитации. Типичным представителем этой группы является трехглавая мышца голени, и особенно камбаловидная. Так, медиальная головка икроножной мышцы содержит 50.8%, латеральная — 50.3%, а камбаловидная мышца — 89% медленных волокон [399].
Основными рецепторными аппаратами, формирующими афферентную информацию о степени сокращения или растяжения мышечных волокон, а также натяжении сухожилия, являются нервно-мышечные веретена и сухожильные рецепторы. Нервно-мышечное веретено расположено в эндомизии параллельно мышечным волокнам. Одним концом оно вплетено в сухожилие, а другим — закреплено в интерстиции мышцы. Под капсулой веретена вдоль него протянуты тонкие интрафузальные мышечные волокна, оплетенные окончаниями чувствительных нервных волокон, которые несут информацию о растяжении мышцы к спинному мозгу. Сухожильный рецептор располагается в сухожилии в непосредственной близости от концов экстрафузальных мышечных волокон. Он представляет собой небольшое веретеновидное образование, в подкапсульном пространстве которого располагаются чувствительные нервные окончания, воспринимающие сокращение мышцы. Кроме альфа-мотонейронов, иннервирующих экстрафузальные (основные) мышечные волокна, имеются гамма-нейроны, иннервирующие интрафузальные волокна, которые обеспечивают регуляцию длины нервно-мышечного веретена и тем самым поддерживают относительное постоянство уровня активности сегментарной рефлекторной деятельности [10].
Cтатья предназначена исключительно для всестороннего информирования о заболевании и о тактике его лечения. Помните, что самолечение может навредить Вашему здоровью. Обратитесь к врачу.
